Timothy M. Perez, PhD
About
Publications
Code
Research
422
Categories
All
(2)
carbon
(1)
misc
(1)
422
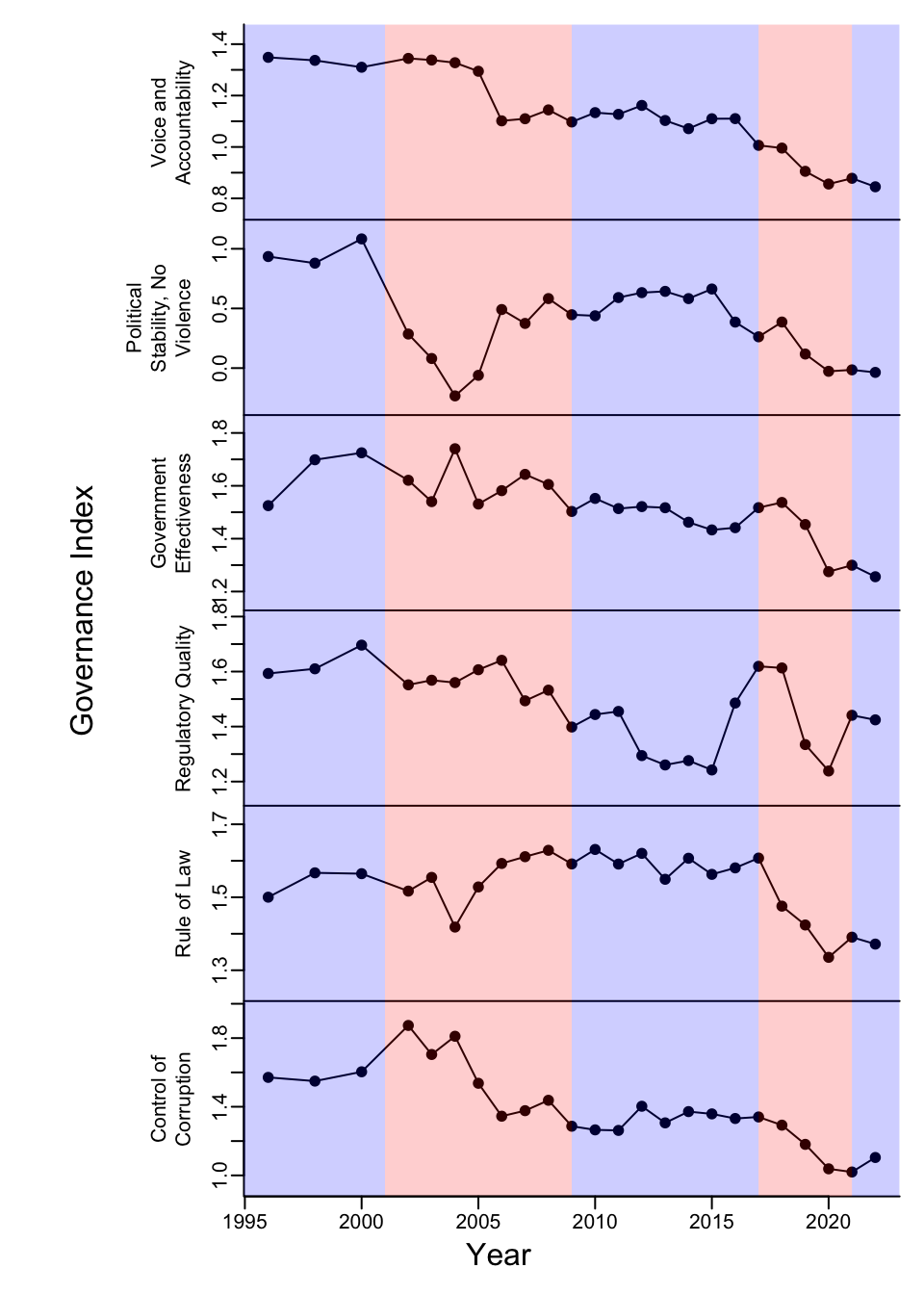
Deteriorating governance increases reversal risk for US AFOLU projects
carbon
Carbon projects reduce or remove greenhouse gasses (GHG) in the atmosphere to mitigate climate change and earn carbon credits. One carbon credit is earned for every metric…
Mar 23, 2024
Timothy Perez
422
misc
This first blog post is mostly intended as a test… but as long as I’m here, I might as well explain why it’s called “422”. This name is relatively arbitrary, I came up with…
Jan 21, 2024
Timothy Perez
No matching items